Результаты и обсуждениеЭкспериментальная генетика и физиология / Территориальное размещение бурых лягушек в период размножения / Результаты и обсуждениеСтраница 2
Как показывают представленные данные, количество жизнеспособной пыльцы зависит у растений от целого ряда факторов (генетических, возрастных, условий среды) и потому значения признака существенно колеблются. В связи с этим были проанализированы не только экспрессия показателя, но и его изменчивость на основе сравнения компонент фенотипиче-ской дисперсии разных по составу комплексов. Для этой цели применен метод двухфакторного дисперсионного анализа, позволяющий оценить влияние генетических факторов и условий среды на процент фертильной пыльцы. У растений мутантных популяций в М1-М5-поколениях специфичность варьирования количества фертиль-ных пыльцевых зерен в значительной степени обусловлена влиянием фактора «поколение» (возрастные особенности мутантных популяций и условия их выращивания в каждом поколении) и составила 61,08% общей фенотипиче-ской дисперсии. Влияние генетического фактора (генетические особенности мутантных популяций) оказалось несущественным (табл. 4). В то же время при различных способах опыления растений влияние специфического генетического фактора (мутантные популяции) на уровень изменчивости процент фертильной пыльцы было значительным.
Таблица 2. Фертильность пыльцы у растений-ревертантов овсяницы луговой на протяжении шести лет культивирования
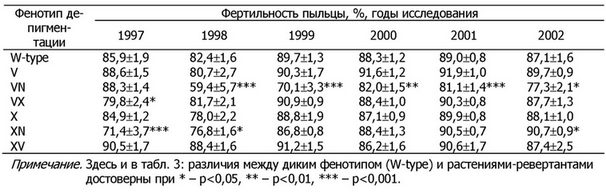
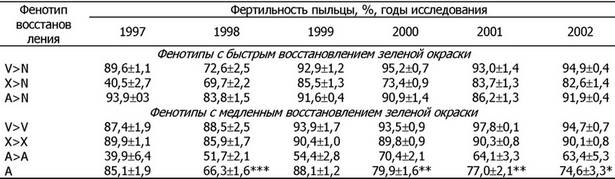
Таблица 3. Фертильность пыльцы у растений-ревертантов овсяницы луговой с различной скоростью реверсии хлорофиллдефектного фенотипа к дикому типу в ювенильной фазе
Так, при гибридизации оно составило 30,92%, при инбридинге — 26,15, а при аут-кроссе — 12.52% от общей дисперсии. Таким образом, при изучении влияния на изменчивость фертильности пыльцы различных факторов необходим анализ разных по составу дисперсионных комплексов, с различным набором и сочетанием специфических (генетических и негенетических) и неспецифических факторов.
У растений-ревертантов другой изученной группы установлено влияние обоих факторов (генотипа и возраста) на уровень изменчивости фертильности пыльцы, но генотипические особенности ревертантов оказались существеннее, чем возраст растений (табл. 4). Последний фактор, как и фактор «поколение» в случае с мутантными популяциями, является неспецифическим, т.к. отражает не только старение растений, но и варьирующие условия их культивирования.
Несомненный интерес как в генетических, так и в селекционных исследованиях представляет оценка действия естественного отбора (стабилизирующей его формы) для признаков, характеризующих жизнеспособность популяций с естественным (супресированная хлорофилл-дефектность) и индуцированным (мутагенез) генетическим грузом. Действие стабилизирующего отбора может быть оценено на основе анализа отклонений от среднего арифметического значения признака (Шмальгаузен, 1969; Алтухов, 2003). Отклонения от среднего значения процента фертильной пыльцы (+ или -) графически представлены прямыми (линии рассеяния, тренды) с разными углами наклона к оси абсцисс, которые отчетливо характеризуют различия как между мутантными популяциями, так и между хлорофиллдефектными фенотипами. Фертильность пыльцы у двух групп мутантных популяций, сформированных на основе действия химических мутагенов и их комбинированного применения с гамма-радиацией, ближе к среднему арифметическому, и это отражается в меньшей крутизне принадлежащих им линейных трендов (рис., А). Напротив, значительно отличается от среднего фертильность пыльцы у контроля и гамма-популяции с соответствующими им крутыми трендами. Паре растений-ревертантов ксанта и виридис также соответствуют пологие линии рассеяния и наименьшие отклонения от среднего значения процента фертильности пыльцы (рис., Б). Альбина-ревертанты и дикий тип имеют более крутые тренды, что связано с большими отклонениями значений от среднего. Если рассматривать эти данные как отражение специфического адаптивного состояния популяций, находящегося под контролем естественного отбора (стабилизирующая форма), то очевидно, что стабилизирующий отбор в большей мере благоприятствует мутантным популяциям, сформированным на основе применения химических мутагенов, и в меньшей — гамма-популяциям. По-видимому, в этом заключается одна из особенностей химического мутагенеза. Ксанта- и виридис-ревертантам стабилизирующий отбор благоприятствует в большей мере, чем альбина-ревертантам, характеризующимся значительными структурными дефектами фотосинтетического аппарата (Венжик и др., 2002) и, как следствие, подвергающимся более жесткому действию естественного отбора. Последние, не являясь летальными, все же частично вносят деструктивный элемент в процессы формирования мик-рогаметофита, что и отражается на количестве фертильных пыльцевых зерен. Следствием таких структурных нарушений может быть и ухудшение жизнеспособности потомства.
Смотрите также
Экономические методы охраны окружающей среды и особенности их использования в России
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...
Экологическая ниша
Понятие ниши пронизывает все сферы экологии. Если бы термину «экологическая ниша»
не придавали так много самых разных значений, то экологию можно было бы определить
как науку о нишах. Многие аспек ...
Взаимодействие климата и растительности
Климат — главный фактор, определяющий характер растительности. Растения в свою
очередь также в некоторой степени воздействуют на климат. Как климат, так и растительность
оказывают решающее влияние ...