Уравнение Лотки — Вольтерра и теория конкуренцииЭволюционная экология / Взаимодействия между популяциями / Уравнение Лотки — Вольтерра и теория конкуренцииСтраница 3
Иногда бывает полезно преобразовать уравнения (1) и (2), помножив выражения, содержащиеся в скобках, на r,N, или на rN соответственно: 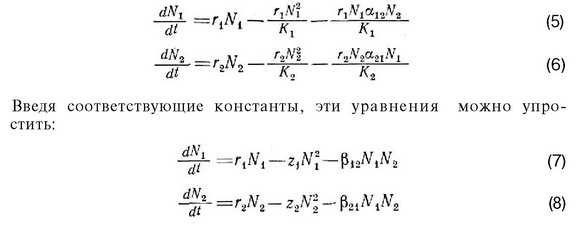
где и 12 соответственно равны г/К, и г/К; а
р21=г2а21. В уравнениях (7) и (8) первый член справа от знака равенства — независимая от плотности скорость увеличения популяции, а второй и третий члены соответственно отражают внутривидовое и межвидовое конкурентное подавление этой скорости увеличения.
Уравнения Лотки — Вольтерра основаны на некоторых допущениях. Часть из них можно сделать менее строгими, но тогда математические манипуляции сразу становятся слишком сложными. Принимается, что максимальные скорости увеличения, коэффициенты конкуренции и предельные плотности насыщения — это неизменные константы, которые не зависят от плотности популяций, состава сообщества и прочих факторов. В результате все эффекты подавления внутри популяций и между ними строго линейны, а все особи N, идентичны, равно как и особи N. Однако к сходным выводам можно прийти и без допущения линейности, используя только графический метод (рис. 6.4). Принимается, что реакции на изменение плотности мгновенны; кроме того, эти два вида не имеют возможности разделиться, т. е. предполагается, что среда соверщенно гомогенна. Были также построены и математические модели конкуренции в пятнистой среде (Skellam, 1947; Levins, Culver, 1971; Horn, MacArthur, 1972; Slatkin, 1974; Levin, 1974). 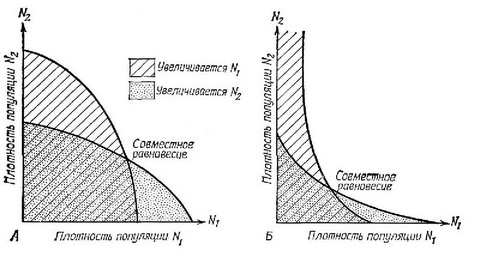
Рис. 6.4. Непрямые изоклины, характеризующие условия устойчивого сосуществования двух конкурентов. Вогнутые изоклины (справа) были нолучены в лабораторных экспериментах на Drosophila (Ayala, Gilpin, Ehrenfeld, 1973), они также присущи некоторым новым моделям конкуренции, например моделям Шёнера (Schoener, 1973, 1976b).
На самом деле в реальных популяциях скорости увеличения, конкурентоспособность и предельные плотности насыщения меняются во времени и пространстве и зависят от свойств особей, плотности популяций и состава сообщества. Действительно, непрерывное изменение среды во времени часто приводит к сосуществованию, которое обусловливается постоянным изменением конкурентоспособности населяющих ее популяций. Эффекты запаздывания (лаг-эффекты), без сомнения, играют в реальных популяциях определенную роль. Наконец, гетерогенность среды может привести к тому, что настоящие конкуренты вырабатывают в процессе дивергенции различные способы использования ресурсов и тем самым ослабляют межвидовое конкурентное подавление.
Почти все теории, построенные на уравнениях конкуренции Лотки — Вольтерра, сталкиваются с вопросом условий равновесия. Но реальные экологические системы (и, следовательно, их части) нередко бывают насыщены не полностью, что само по себе может привести к сосуществованию популяций, которые в иных условиях не выдерживают конкуренции. Например, понижение уровня плотности популяций и связанное с этим ослабление конкуренции могут произойти в результате независимого от плотности разрежения или хищничества. Однако введение в уравнение Лотки — Вольтерра фактора разрежения, ведущего к понижению плотности популяций, не приводит к ослаблению интенсивности конкурентного давления. Более реалистичная модель должна учитывать отношение мгновенной потребности к снабжению. Например, можно считать, что коэффициенты конкуренции являются переменными, зависящими от общей плотности обеих популяций.
На уровне насыщения (К,, К или + на рис. 6.3, Г) отношение потребность/снабжение равно единице, а коэффициенты конкуренции цринимают максимальные значения, но но мере достижения йодного конкурентного вакуума они будут стремиться к нулю.
Уравнения конкуренции Лотки — Вольтерра играют чрезвычайно важную роль в современной экологической теории (Levins, 1968; MacArthur, 1968, 1972; Vandermeer, 1970, 1972; May, 1976), однако большое число содержащихся в них нереалистичных с биологической точки зрения допущений расходится с существующей теорией
Смотрите также
Взаимодействие климата и растительности
Климат — главный фактор, определяющий характер растительности. Растения в свою
очередь также в некоторой степени воздействуют на климат. Как климат, так и растительность
оказывают решающее влияние ...
Особенности природоохранной деятельности в России.
В нашей стране на первом этапе становления хозяйственного
механизма природопользования недостатки административной системы руководства
проявлялись более явно и отчетливо, чем в других странах. ...
Роль изоферментов лактатдегидрогеназы в адаптациях млекопитающих Карелии
В экстрактах тканей сердца, почек, скелетных мышц, печени, легких, селезенки
у американской норки (Mustela vison L.), песца (Aiopex iagopus), лисицы ( Vuipes
vuipes L.), лесной куницы (Martes mart ...