Представления континуалистов об ареалеЭкология морского бентоса / Биогеография — общие вопросы / Представления континуалистов об ареалеСтраница 3
Значения абиотических условий, ограничивающих распространение вида, естественно, различны для разных видов, поэтому границы потенциальных ареалов отдельных видов могут совпадать только случайно. Если бы градиент абиотических условий на поверхности Земли был бы постоянен, то границы ареалов видов распределялись бы случайно. В действительности наблюдается чередование областей с резкими и слабыми градиентами. Области резких градиентов являются аттракторами границ (см. предыдущую главу), поэтому на поверхности Земли чередуются области сгущения и разрежения границ ареалов. Это чередование можно обнаружить статистическими методами. Континуалист-ская биогеография считает, что причиной появления областей сгущения границ ареалов является наличие областей с резкими градиентами абиотических факторов, и свою основную задачу видит в выявлении таких областей.
Я.И. Старобогатов (1970: 91) писал: «Фактические ареалы будут тем меньше потенциальных, чем большее значение в распространении данного вида играют исторические факторы», — под которыми он понимает процесс расселения вида из места его возникновения. «Районирование, построенное на легко расселяющихся организмах, будет менее дробным, чем построенное на организмах, обладающих малыми способностями к расселению» (Старобогатов, 1982: 16). Разбирая эту умозрительную гипотезу, прежде всего надо отметить, что предположение о прямой связи размера ареала с лёгкостью расселения отнюдь невсегда подтверждается фактическими данными.
Представление о влиянии малых способностей к расселению на размер ареала основано на смешении двух совершенно разных явлений: способности к расселению самой по себе и способности к преодолению преград. В качестве примера рассмотрим распространение полихетыAnobothruspatersoni (рис. 5.6). Хотя биология размножения этого вида неизвестна, но он принадлежит к семейству, все изученные виды которого не имеют пелагических личинок, поэтому с высокой вероятностью можно считать, что пелагической личинки нет и у него. Данный вид — стенобатный абиссальный, известен с глубин 3460-8050 м, и потому проникновение его в Северную Атлантику из Северной Пацифики через Северный Ледовитый океан исключено. Этот ареал и многочисленные аналогичные примеры однозначно свидетельствуют об отсутствии связи размера ареала вида с его способностью к расселению в пределах ЦС, в которые он входит. Вероятно, это можно объяснить тем, что за время существования ЦС любой вид со сколь угодно малыми способностями к расселению способен расселиться по всей её площади. Другое дело, если вид в своём распространении встречает преграды. В подобном случае способность к их преодолению действительно зависит от способности вида к расселению. Многочисленные примеры данной связи можно найти в ареалах пресноводных беспозвоночных и биогеографии островов. 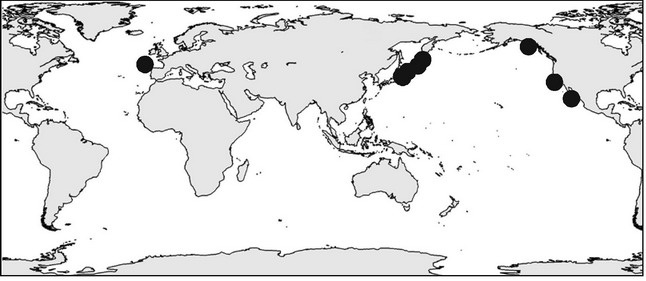
Рнс. 5.6.
Ареал Anobothrus patersoni по И.А. Жирков (2009). 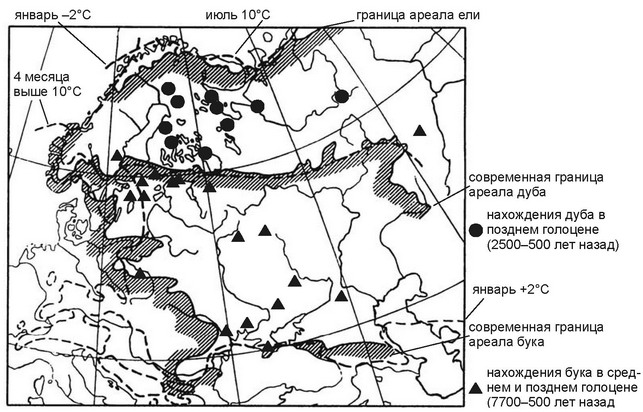
Рис. 5.7.
Северные границы ареалов ели Picea abies и дуба Quercus robur и восточная — бука Fagus sylvatica в сопоставлении с клималиниями (прерывистые линии). По Г. Вальтеру (1982). Нахождения дуба и бука. По С.А. Турубановой (2000) и О.В. Смирновой и С.А. Турубановой (2004).
Смотрите также
Платежные инструменты
Платежные инструменты - это правовые средства осуществления переводов денежных средств (наличных и безналичных) с целью совершения платежа.
Порядок выполнения операций с применением платежных инст ...
Экологическая ниша
Понятие ниши пронизывает все сферы экологии. Если бы термину «экологическая ниша»
не придавали так много самых разных значений, то экологию можно было бы определить
как науку о нишах. Многие аспек ...
Биогеография
Основная цель экологии состоит в том, чтобы понять те разнообразные факторы,
которые влияют на распределение и обилие животных и растений (Andrewartha, Birch,
1954; Krebs, 1972; Mac-Arthur, 1972). ...