Затраты на потомствоЭволюционная экология / Принципы популяционной экологии / Затраты на потомствоСтраница 2
Молодые и взрослые особи часто испытывают очень разное давление отбора. Репродуктивное усилие должно отражать действие факторов внешней среды на взрослых, тогда как затраты на потомство находятся под сильным влиянием той среды, в которой сушествует молодь. Поскольку любые две части триумвирата определяют третью, оптимальный размер кладки или потомства является прямым следствием оптимального текушего репродуктивного усилия в сочетании с оптимальными затратами на потомство (размер кладки фактически равен отношению репродуктивного усилия к затратам на одного потомка). Конечно, размер кладки сам по себе подвержен действию естественного отбора. Вспомним пример рогатых яшерип, которые живут долго и во взрослом состоянии подвержены _ЙГ-отбору, но которые благодаря своей бо-чонкообразной форме тела характеризуются значительным репродуктивным усилием и продупируют множество мелких потомков, подверженных очень высокой смертности (см. с. 106—108).
Величина кладки у птиц
в настояшее время собран богатый материал, касающийся оптимальной величины кладки у птип (Lack, 1954, 1966, 1968). Этими изящными исследованиями показано, что кладки средней величины в сравнении с очень малыми или очень большими дают (про-порпионально своей величине) больше потомков, доживающих до момента собственного размножения в следующем сезоне. Данное наблюдение является прекрасным примером стабилизирующего отбора. Птеппы, вылупившиеся в больших кладках, оставляя гнездо, характеризуются меньшим весом (рис. 5.22) и значительно более низкой выживаемостью в послегнездовой период. Очевидно, оптимальное число яип в кладке такое, при котором родители в среднем способны обеспечить достаточным количеством пищи всех 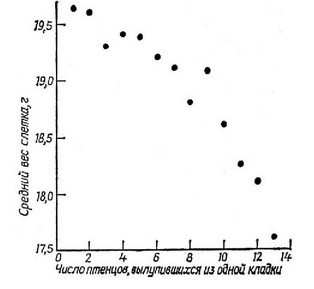
Рис. 5.22. Зависимость среднего веса слетка большой синицы от величины кладки; видно, что вес молодых особей в больших кладках снижается.
Рис. 5.23. Средняя величина кладки в популяции большой синицы, меняющаяся на протяжении 17 лет в зависимости от плотности гусениц. Заметна тенденция к увеличению размера кладки в годы, когда гусеницы имеются в изобилии.
птенцов. Это положение хорошо иллюстрируется (Perrins, 1965) данными по изучению популяции большой синицы (Pams major), величина кладки у которой на протяжении 17-летнего периода варьировала от 8 до 12 яиц, что зависело от количества основной пищи — гусениц (рис. 5.23). Винн-Эдвардс (Wynne-Edwards, 1962) считает, что оптимальна такая кладка, при которой численность молодых, достигших периода размножения, будет точно равна численности родителей. Подобное объяснение само по себе включает понятие группового отбора (см. гл. 1), поскольку отдельные птицы не обязательно стремятся иметь как можно больше потомков, а производят их ровно столько, чтобы заменить самих себя. Ясно, что «мошенник», дающий больше потомков, должен вскоре «затопить» генофонд своими генами. В конце главы мы еще вернемся к этому вопросу.
Даже в пределах одного распространенного вида птиц или млекопитающих особи, живущие в более высоких широтах, производят больше потомков, чем живущие в более низких широтах (рис. 5.24). Явление увеличения кладки (помета) в зависимости от географической широты весьма распространено, благодаря чему 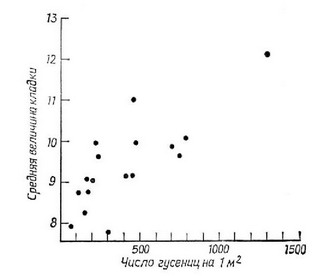
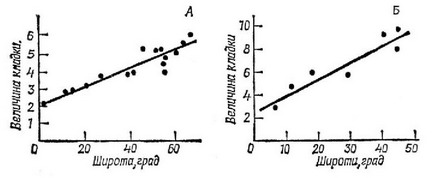
Рис. 5.24. Зависимоств размера кладки от географической щироты для птиц рода Emberiza (А) и Охуига (Б). (Из Cody, 1966.)
оно и привлекло внимание многих популяционных экологов. Для объяснения этого явления предложено несколько гипотез, отнюдь не взаимоисключающих друг друга.
Гипотеза продолжительности светового дня. Как указывалось в гл. 2, в конце весны и летом продолжительность дня в высоких широтах больше, чем в низких. Вследствие этого дневные птицы располагают большим светлым временем суток для сбора пищи и, таким образом, способны прокормить больше птенцов. Однако у птиц и млекопитающих, ведущих ночной образ жизни, величина кладки (или помета) также возрастает с широтой, хотя очевидно, что виды эти имеют более короткий период для сбора пищи.
Гипотеза весеннего изобилия (или гипотеза конкуренции). Многие птицы умеренной зоны совершают перелеты, тогда как в тропиках таких видов очень мало. Весной в средних широтах наблюдается резкое увеличение первичной продукции и быстрое возрастание численности насекомых, зависящих от этого источника вещества и энергии. Численность же популяций самих птиц, как перелетных, так и оседлых, весной очень невелика из-за массовой гибели в зимний период. Поэтому прилетающие весной особи оказываются в условиях обилия пищи и сравнительно слабой конкуренции. Зимующие в тропиках перелетные виды испытывают там обычную для этих мест острую конкуренцию, тогда как в умеренной зоне в весенние месяцы конкуренция значительно ослабляется. Таким образом, птицы более высоких широт благодаря тому, что они могут собрать больше корма в единицу времени, способны прокормить большее число потомков до того возраста, когда последние начнут питаться самостоятельно.
Смотрите также
Платежные инструменты
Платежные инструменты - это правовые средства осуществления переводов денежных средств (наличных и безналичных) с целью совершения платежа.
Порядок выполнения операций с применением платежных инст ...
Понятие исследования
В современном менеджменте исследование является главным фактором успеха, а если выражаться по научному, — главным фактором повышения эффективности управления. Исследование — это не только ...
Дно как биотоп
Люди довольно хорошо представляют себе, как выглядит поверхность суши. Однако
о том, как выглядит поверхность 3/4 Земли — морское дно и какие там условия, у подавляющего
большинства представления ...