Классификации, рассматривающие три жизненные стратегииЭкология морского бентоса / Аутоэкология / Классификации, рассматривающие три жизненные стратегииСтраница 2
G.J. Vermeij (1978) считает, что логическими основаниями для выделения трёх типов стратегий являются особенности окружающей среды (судя по тексту — благоприятность условий, в частности, их стабильность) и то, как виды реагируют на них. Он полагает, что для видов с высокой скоростью роста популяции, которых он называет не эксплеренты, а оппортунисты
(т. е. считает их синонимом r-стратегов), характерна высокая способность к колонизации эфемерных или часто нарушаемых биотопов благодаря высокой способности к расселению. Однако их конкурентоспособность низка, поэтому виды других групп вытесняют их. Для патиентов
характерна способность длительно существовать в неблагоприятных условиях (выносить физиологический стресс). Их отличает низкая скорость роста и, независимо от условий их существования, невысокий репродуктивный потенциал. В результате при благоприятных условиях, они также растут очень медленно, поэтому их конкурентоспособность низка. В качестве примера таких видов он приводит виды супралиторали и верхних горизонтов литорали, абиссали и районов с пониженной солёностью. Многие черты К-стратегов возможно связаны не с их высокой конкурентоспособностью, а с устойчивостью особей таких видов, поэтому К-стратеги и патиенты могут иметь немало сходного в своей биологии. К третьей группе он относит биотически успешные виды,
обитающие в условиях, близких к оптимальным. В эту группу попадают не только К-стратеги. Если К-стратеги по определению поддерживают плотность своих популяций на высоком уровне благодаря конкурентному превосходству во взаимодействии с другими видами, то биотическая успешность видов, по мнению G.J. Vermeij, может иметь и другие причины. Биотически успешные виды также имеют большую продолжительность жизни, сравнительно высокую скорость роста на ранних стадиях развития. В море, по его мнению, такие виды преобладают на наружной поверхности рифов, в верхних нескольких сантиметрах рыхлых грунтов мелководий, в нижней литорали. 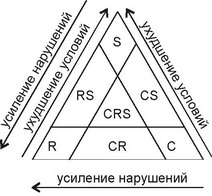
Рис. 2.23.
Треугольник Грайма.
С, К, 8 — основные типы жизненных стратегий, СК, С8, СК8 — переходные между ними типы жизненных стратегий. По Миркин и др. (2001).
Многие живые организмы имеют короткую продолжительность жизни и не имеют выраженного периода индивидуального роста или, по крайней мере, не имеют выражен-ной возрастной структуры популяции (простейшие, фитопланктон, многие виды зоопланктона, особенно пресноводного и др.), поэтому к ним не применимы логические основания классификаций, использованные Л.Г. Раменским, L. Van Valen, J.P. Grime и G.J. Vermeij.
Ю.Э. Романовский (1989а,б), изучавший пресноводный зоопланктон, обратил внимание на то, что в природе обычно ресурсы испытывают флуктуации и в качестве логического основания предложил использовать амплитуду и частоту колебания ресурса. По его мнению, при больших амплитудах колебания ресурса высокой численности популяции достигают r-стратеги (ярко выраженные эксплеренты), способные быстро увеличить численность. При низкой амплитуде высокой численности популяции достигают К-стратеги (крайние патиенты), которые не только сами устойчивы к минимальной концентрации ресурса, но и «поддерживают его количество на постоянном и низком уровне, недоступном для других видов» (1989а: 25). С другой стороны, продолжительность периодов дефицита ресурса также различна. Если период дефицита ресурса существенно превышает продолжительности жизни особей, то такие r-стратеги (эксплеренты) переживают этот период на латентной стадии (споры и семена некоторых растений, покоящиеся яйца беспозвоночных, а также особи, способные впадать в состояние анабиоза). Если период дефицита ресурса меньше продолжительности жизни особей, то такие r-стратеги (виоленты) переживают его на половозрелой стадии. Таким образом, С-S-R-стратегии — это адаптации к разным уровням колебания ресурсов независимо от природы таких колебаний, и положение вида в С-S-R-пространстве определяет распределение расхода энергии на различных стадиях онтогенеза.
Л.В. Ильяш (Ильяш и др., 2003) на основе анализа популяционных стратегий фитопланктона в качестве логических оснований классификации стратегий предложила использовать способность вида конкурентно воздействовать на совместно обитающие с ним виды и чувствительность к такому воздействию со стороны других видов. При увеличении значений одного показателя уменьшается другой и наоборот. Эксплеренты (R-страте-гия) и виоленты (С-стратегия) занимают при этом крайние положения, патиенты (S-стра-тегия) — среднее, одновременно только для них характерна высокая устойчивость к низкой концентрации ресурса (рис. 2.24).
Смотрите также
Платежные инструменты
Платежные инструменты - это правовые средства осуществления переводов денежных средств (наличных и безналичных) с целью совершения платежа.
Порядок выполнения операций с применением платежных инст ...
Экономические механизмы охраны природы
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...
Основные функции и принципы экологической политики.
Комплексный характер экологических проблем
требует комплексного государственного управления в области охраны окружающей
среды. Ниже перечислим функции такого управления.
* Экологическое
прогно ...