Практика и традиции выделения сообществ макробентосаЭкология морского бентоса / Сообщества / Практика и традиции выделения сообществ макробентосаСтраница 1
Дно, в отличие от поверхности суши, длительное время было недоступно для визуального наблюдения. Изучение бентоса с судна традиционными орудиями лова можно сравнить с изучением суши с воздушного шара, плывущего над облаками. Кто-то сказал, что с помощью трала в этом случае можно собрать мартовских котов и пару трубочистов. Исследователь-гидробиолог, в отличие от коллег, занимающихся изучением наземных экосистем, не имеет возможности отбирать пробы в конкретном сообществе, он может ориентироваться лишь весьма приблизительно, основываясь на данных по рельефу и материалам предыдущих съёмок.
Первые исследования при описании бентоса выделяли не сообщества, а описывали отличия биоты разных грунтов. Например, в классическом труде по бентосу Чёрного моря, С.А. Зёрнов (1913) выделял биоценозы скал, песка, ракушечника, илистых берегов, мидиевого и фазеолинового ила и т. д. Такая практика выделения «сообществ» встречается и сейчас.
Постепенно было обнаружено, что в конкретном месте один из видов резко доминирует над остальными. Станции с доминированием одного вида стали относить к одному сообществу. Особенно широкое распространение практика выделения сообществ по видам-доминантам получила в отечественной гидробиологии (Броцкая, Зенкевич, 1939; Шо-рыгин, 1955 и др.). Этот метод выделения сообществ в изучении бентоса, как и в некоторых школах геоботаников, сохранилась до сих пор.
В исследованиях бентоса доминирование вида определяют в конкретной пробе или группе проб, взятых на одной и той же или соседних станциях (Броцкая и Зенкевич, 1939; Воробьев, 1949; Нейман, 1963). Доминирование следует оценивать по роли, которую играет вид в сообществе. Длительное время в качестве такого показателя использовали долю биомассы часто с учётом частоты встречаемости. За рубежом нередко вместо биомассы используют численность. Однако оба этих показателя имеют недостатки. При выделе-нии доминантов по биомассе таковыми становятся крупные, но медленно растущие моллюски и иглокожие. При использовании численности в том же сообществе доминантом
Таблица 3.1.
Средние значения коэффициента удельной интенсивности метаболизма (с) нри стандартной темнературе 20°С, алометрического ноказателя и калорийности для некоторых грунн макрозообентоса. По Алимову (1979); Кучеруку (1985); Голикову и др. (1988). 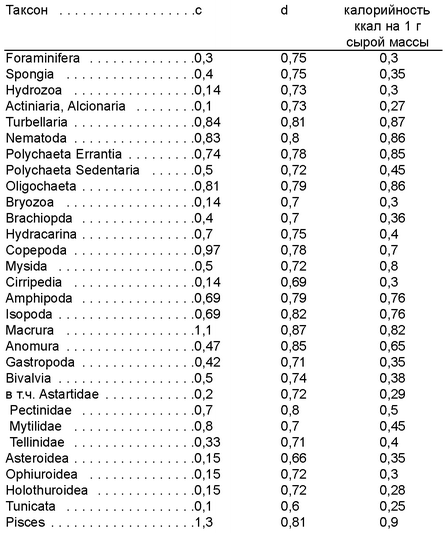
могут стать мелкие полихеты, имеющие низкую биомассу. Поскольку сообщество — функционирующая структура, правильнее для выделения доминирующих видов применить не структурные показатели (численность, биомассу), а функциональные. В качестве такого показателя в последнее время широко используют поток энергии через популяции видов. Поток энергии можно считать пропорциональным интенсивности метаболизма популяции (трат на обмен). Интенсивность метаболизма популяции рассчитывают для животных по формуле: 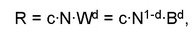
где W — средний индивидуальный сырой вес особи, N — численность и В — биомасса на 1 м2, d — аллометрический показатель, с — специфичный для таксона коэффициент удельной интенсивности метаболизма.
При расчете потока энергии для макробентоса величину d обычно принимают равной 0,75, можно использовать значения из табл. 3.1; значения с специфичны для разных таксонов (табл. 3.1). В этой же таблице приведены средние величины удельной калорийности в ккал на 1 г сырой массы.
При оценке обилия раковинных моллюсков в значения сырого веса обычно вводят поправки на среднюю весовую долю их раковины. Этот показатель колеблется в весьма широких пределах в зависимости от таксона, и его лучше определить самому для конкретного материала. В качестве первого приближения можно использовать значения для Gastropoda — 0,5-0,67, Bivalvia — 0,9.
Анализ имеющихся в литературе данных о продуктивности различных групп одноклеточных водорослей (морских и пресноводных), в основном диатомовых и перидиние-вых, показывает, что значения коэффициента d для этих организмов близки к 0,5 (Гутель-махер и др., 1980; Гутельмахер, 1983; Заика, 1983; Raven, Kubler, 2002).
Смотрите также
Экологическая ниша
Понятие ниши пронизывает все сферы экологии. Если бы термину «экологическая ниша»
не придавали так много самых разных значений, то экологию можно было бы определить
как науку о нишах. Многие аспек ...
К популяционной организации политипического вида (на примере рыжей полевки -
clethrionomys glareolus shreb.)
Анализ популяционной организации и динамики численности европейской рыжей полевки
- типичного представителя мегаареальных политипических видов млекопитающих Палеарктики
- подтвердил высказанное на ...
Морфофункциональные особенности лейкоцитов млекопитающих, разводимых в неволе
в условиях европейского севера
Представлены данные о морфофункциональных особенностях лейкоцитов крови различных
видов животных из отряда Carnivora— норок, песцов, лисиц и енотовидных собак, разводимых
в неволе в условиях Карел ...