Сукцессии и взаимоотношения макрофитовЭкология морского бентоса / Сообщества / Сукцессии и взаимоотношения макрофитовСтраница 1
При освоении нового субстрата растительность проходит несколько сукцессионных стадий (рис. 3.44). Первыми (в течение суток) твёрдый субстрат заселяют диатомеи, при чём как бентосные, так и планктонные. Последние, однако, через неделю исчезают из сообщества. Для этой стадии характерно очень высокое видовое разнообразие. Через 9-12 дней появляются первые макрофиты. Все они — высокопродуктивные быстрорастущие оппортунисты с нитчатыми или трубчатыми талломами: зелёные Ulothrix, Enteromorpha spp., красные Ceramium spp., Polysiphonia и др. Развиваются и вертикальные колонии ди-атомей (Navicula). Эта стадия сукцессии длится 30-120 дней от начала заселения. Её называют фазой ульвовых, поскольку преобладающими в ней являются ульвовые зелёные водоросли. В переходном периоде между первой и второй фазами видовое разнообразие диатомей падает, их покров из сплошного становится пятнистым, и постепенно первые «колонизаторы» исчезают. Третья фаза (нормально структурированного сообщества) характеризуется появлением и развитием многолетних форм с меньшими скоростями роста. Она начинается примерно через 3 месяца. Изменения во флористическом составе сообщества сопровождают существенные функциональные изменения при развитии сукцессии: видовое разнообразие падает к концу каждой фазы и возрастает в начале следующей при общем тренде увеличения; коэффициент P/B стремительно падает от первой фазы к третьей; концентрация хлорофилла а, напротив, существенно возрастает. Показано, что в переходные моменты от одной фазы к другой увеличивается поток энергии через сообщество, а рассеяние энергии последовательно уменьшается от первой фазы к третьей. При этом растительность третьей фазы зависит от степени развития растительности второй, в то время как фаза ульвовых не зависит от первой фазы, и интенсивность её развития определяется только стратегиями роста её компонентов (Niel, Valera, 1984). Таким образом, взаимовлияние макрофитов начинает сказываться на ранних стадиях сукцессии. Seo local seo.
Структуру фитоценозов определяют биотические взаимодействия и абиотические факторы. Биотические взаимодействия могут быть как в пределах фитоценоза (конкуренция за субстрат, свет, биогены и т. д.), так и между растениями и остальными компонентами сообщества. 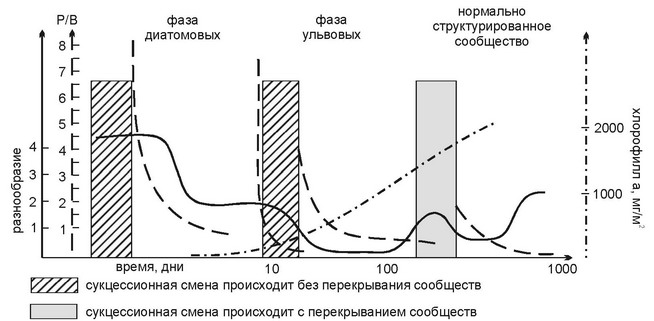
Рис. 3.44.
Сукцессия водорослей на новом субстрате 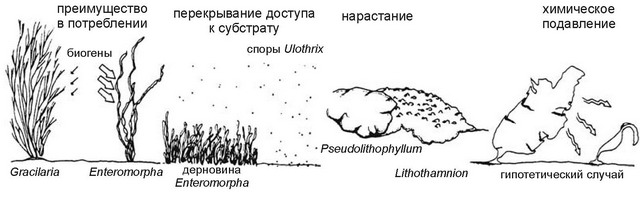
Рис. 3.45.
Механизмы реализации конкуренции. По Olson, Lubchenco (1990).
Конкуренция влияет на возрастную и размерную структуру, а также на успех размножения отдельных особей. Конкурентоспособность макроводорослей определяется всем комплексом их характерных черт как морфологических (форма и степень разветв-лённости таллома, его толщина и прочность), так и физиологических (чувствительность к свету, способы поглощения и запасания питательных веществ, выделение защитных химических соединений, способность к модификациям) (Olson, Lubchenco, 1990).
Можно выделить несколько механизмов реализации конкурентных способностей (рис. 3.45): способность к интенсивному потреблению питательных веществ; способность к быстрому захвату субстрата с образованием плотных зарослей-дерновин (turf); способность к непосредственной топической конкуренции — т. е. к механическому вытеснению конкурентов через нарастание на их талломы; аллелохимическое воздействие на конкурентов (через антибиотики и токсины).
Первый механизм иллюстрирует взаимоотношения зелёной ульвовой Enteromorpha sp. и красной Gracilaria tikvahiae. Если содержание питательных веществ в среде постоянно, то r-стратег энтероморфа вытесняет грацилярию за счёт существенно более высокой скорости потребления. Но грацилярия имеет более высокую запасающую способность, поэтому при значительных колебаниях концентрации питательных веществ она получает конкурентное преимущество и вытесняет энтероморфу.
Второй механизм свойственен дерновино-образующим водорослям. На рис. 3.45 представлена ситуация, при которой споры Ulothrix просто не имеют возможности достичь субстрата, сплошь покрытого дерновиной энтероморфы. Аналогичная картина наблюдается во взаимоотношениях энтероморфы с зиготами Fucus vesiculosus. Однако, при появлении фитофагов ситуация меняется: фитофаги выедают пятна в покрове энтероморфы, зиготы фукуса оседают в них, и в течение некоторого времени фукус вытесняет энтероморфу, которая полностью исчезает под его покровом, но может перейти к эпифитиро-ванию на выросших талломах фукуса. Если же удалить фитофагов после оседания зигот фукуса, то энтероморфа довольно быстро «забьёт» проростки фукуса, закрыв их от света. Т. е. при определении конкурентных отношений между видами следует учитывать различную конкурентную способность разных онтогенетических стадий.
Смотрите также
Особенности природоохранной деятельности в России.
В нашей стране на первом этапе становления хозяйственного
механизма природопользования недостатки административной системы руководства
проявлялись более явно и отчетливо, чем в других странах. ...
Структура сообщества
За исключением краткого рассмотрения биомов в гл. 3, до сих пор мы касались только
экологии особей и популяций. В этой главе речь пойдет об экологии сообществ. Подобно
тому как популяции обладают ...
Влияние окружающей среды на живые организмы
Все процессы в биосфере взаимосвязаны. Человечество - лишь незначительная часть биосферы, а человек является л ...