Формирование травяного биомаЭкология морского бентоса / Синэкология / Формирование травяного биомаСтраница 1
В качестве примера возникновения ЦС путём преобразования рассмотрим возникновение травяных ЦС (ТЦС) — степей, прерий, пампасов, саванн и др. Их формирование обычно связывают с дефицитом влаги (Мордкович, 1982). С.М. Разумовский (1999), проанализировав данные по связи распределения растительности с температурой и влажностью в 531 точках Земного шара, обнаружил, что распределение климаксных фитоценозов не связано с влажностью, что объяснил способностью сукцессионных систем эффективно регулировать влажность своих биотопов.
С другой стороны, палеонтологические данные свидетельствуют об отсутствии безлесных ЦС в раннем палеогене (палеоцене и начале эоцена). В то время климаксными ассоциациями повсеместно являлись лесные несмотря на то, что на Земле, несомненно, и тогда были районы с теми же условиями, в которых ныне существуют травяные ЦС.
Эти и другие факты заставляют искать иные причины возникновения и широкого распространения безлесных ЦС. Наиболее аргументированной является теория В.В. Жерихина (1993, 1994, 2003), основные положения которой изложены ниже.
Имеющиеся палеонтологические данные свидетельствуют о том, что ТЦС формировались неоднократно и независимо, но всегда сходным образом. Сначала появлялись крупные листоядные позвоночные, которые замедляли прохождение сукцессий в местах, где появление лесных сукцессионных стадий тормозилось климатически, в первую очередь, из-за снижения количества осадков. В результате площадь, занятая травяными сообществами, увеличивалась, что в свою очередь вызывало появление разнообразной травоядной фауны, ещё более тормозившей прохождение сукцессий. Процесс, таким образом, развивался с положительной обратной связью. В конечном итоге формировались эндогенно стабилизированные травяные сообщества с высоким уровнем диффузных коадантаци-онных связей, весьма лабильных и обеспечивающих гибкое регулирование функционирования ассоциаций в целом в условиях изменчивой среды (т. е. ЦС пастбищного типа). В этих сообществах развивались специфичные для них блоки копрофагов, травоядных насекомых (прямокрылых, термитов), санрофагов, некрофагов и др. Доминирующими продуцентами стали травянистые растения, адаптированные к быстрому росту после стравливания. Везде для ТЦС характерны очень высокая продуктивность и гораздо более плодородные почвы, чем в соседних лесных ЦС. 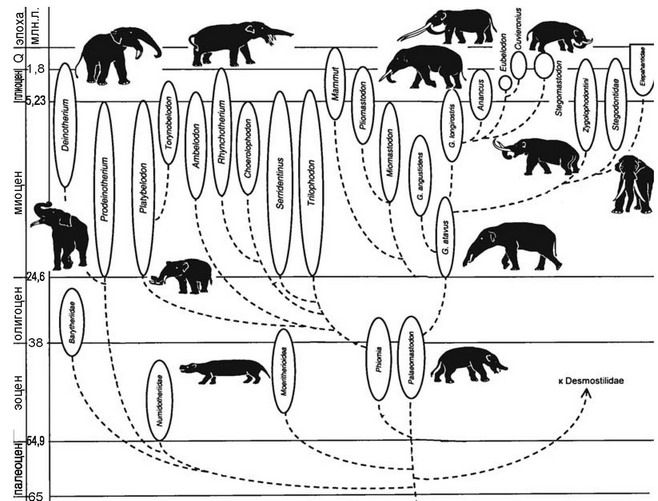
Рис. 4.41
Филогенетическое древо хоботных. По А.К. Агаджаняну (2004).
Когда впервые возникли ТЦС неизвестно. Вполне можно допустить их формирование на основе папортникообразных. Первые достоверно известные ТЦС отмечены в позднем мелу. Их эдификаторами стали растительноядные динозавры (в первую очередь цера-топсиды). Травяной блок был образован не злаками, как ныне, поэтому их нельзя рассматривать как предшественников современных. По неустановленным причинам меловые ТЦС бесследно исчезли.
В кайнозое ТЦС сформировались на основе злаков, причём независимо, как минимум в трёх местах. Первые по времени (в эоцене) ТЦС сформировались в Южной Америке. Их эдификаторами были неполнозубые и вымершие отряды южноамериканских копытных — ксенунгуляты (Xenungulata) и нотоунгуляты (Notoungulata), а позднее — и крупные грызуны, парнокопытные, мозоленогие и хоботные. В Северной Америке ТЦС начинают формироваться в олигоцене, а в миоцене этот процесс захватывает и Евразию с Африкой. Эдификаторами этих ТЦС являлись и остались хоботные и парно- и непарнокопытные. И, наконец, в неогене ТЦС начали формироваться в Австралии. Их эдификаторами были травоядные сумчатые и крупные птицы. В Африке, Азии и Австралии существенную роль в ТЦС играли и разные неродственные таксоны гигантских (до 3 м длиной) черепах, в Южной Америке их, по-видимому, заменяли сходные внешне гигантские броненосцы. Даже в Европе до начала периода оледенений жила черепаха Cheirogaster рег-р1п1апа, достигавшая более 2 м длины. В Новой Зеландии туссоковые злаковники, вероятно, поддерживали страусы моа и иные крупные бескрылые птицы. Возможно, ТЦС формировались и в других местах: на Мадагаскаре (эдификаторы — страусы эпиорнисы).
Процесс формирования травяных биомов и эволюции таксонов-эдификаторов шёл параллельно. Так, хоботные дали вспышки таксономического разнообразия в начале олигоцена, в миоцене, в первой половине плиоцена (рис. 4.41) — когда травяные биомы начали образовываться. После их становления разнообразие хоботных снизилось. Хоботные не просто осваивали новую адаптивную зону, они её формировали.
О ведущей роли крупных травоядных в возникновении ТЦС свидетельствует также то, что после их уничтожения исчезают и ТЦС. Весьма показательно, что голоценовое вымирание крупных травоядных, являвшихся эдификаторам
Смотрите также
Платежные инструменты
Платежные инструменты - это правовые средства осуществления переводов денежных средств (наличных и безналичных) с целью совершения платежа.
Порядок выполнения операций с применением платежных инст ...
Структура сообщества
За исключением краткого рассмотрения биомов в гл. 3, до сих пор мы касались только
экологии особей и популяций. В этой главе речь пойдет об экологии сообществ. Подобно
тому как популяции обладают ...
Биогеография
Основная цель экологии состоит в том, чтобы понять те разнообразные факторы,
которые влияют на распределение и обилие животных и растений (Andrewartha, Birch,
1954; Krebs, 1972; Mac-Arthur, 1972). ...