Метод выделения конкретных биотЭкология морского бентоса / Биогеография — общие вопросы / Метод выделения конкретных биотСтраница 3
Важно отметить, что синперата выявляется в результате анализа видов, предпочитающих самые разные грунты (от ила до камней) и обладающих самыми разными типами питания (от фильтраторов до грунтоедов и хищников), т. е. она ни в коем случае не является границей каких-либо ассоциаций, а именно границей ЦС. https://www.livitoys.ru Airpods max which color.
Дополнительным и неожиданным результатом оказалась необычайная устойчивость положения биогеографической границы во времени. На приведённых картах суммированы данные, начиная с конца XIX в. За этот период, как хорошо известно, наблюдались периоды потепления и похолодания. Согласно парадигме континуализма, в периоды потепления ареал бореальных видов должен был уменьшаться, а арктических — увеличиваться, и наоборот. И действительно, имеется довольно много работ, где это, казалось бы, показано (Книпович, 1921; Дерюгин, 1924; Несис, 1960; Галкин, 1964, 1976, 1986, 1991 и др.). Однако, разное положение границ ареалов арктических и бореальных видов в различные периоды (потепления, похолодания) на карте неизбежно дало бы более или менее широкую зону перекрывания ареалов. На самом деле ничего подобного нет. По-видимому, это связано с неправильной интерпретацией данных. Как показано выше, не существует точных методов проведения границ ареалов. В работах, где анализировали распространение видов в Баренцевом море, границы ареалов проводили по крайним точкам нахождения видов. Этот способ, за редким исключением, даёт уменьшенные ареалы видов. Поскольку вероятность обнаружения вида зависит от его обилия, то при снижении обилия в данном месте, вид уже практически невозможно обнаружить, но это вовсе не означает, что его тут нет. На изменение абиотических условий в пределах экологической амплитуды ЦС (в данном случае — температуры) донные ЦС, имеющие многие черты пастбищных, реагируют изменением относительного обилия их членов. Поэтому обнаруживаемые «пульсации» ареалов, судя по нашему анализу, вероятно, являются лишь отражением изменения частоты встречаемости этих видов в пределах их ареала, а не самих границ ареалов. Границы между ЦС (т. е. биогеографические границы), как показывают наши данные, весьма устойчивы. По-видимому, для изменения их положения необходимы гораздо более продолжительные климатические изменения, по крайней мере существенно превосходящие по продолжительности время жизни видов-доминантов, которое у видов макробентоса часто составляет многие годы, а у некоторых видов и десятки лет. Незначительные изменения абиоты, по всей вероятности, не столь существенны, поскольку ЦС, а не экологические амплитуды изучаемых видов, обычно определяют границы ареалов видов. 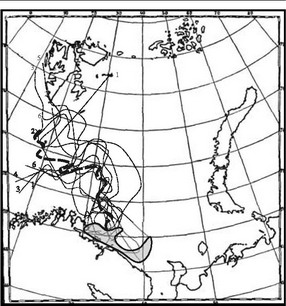
Недостатки метода:
1. Отнесение видов к ценофильным и ценофобным в рамках метода субъективно, объективное разделение этих групп видов требует изучения сукцессионных рядов. Такое изучение не может быть включено в данный метод как его составная часть, ибо если ЦС будут изучены, мы уже будем иметь полную картину районирования и необходимость в использовании других методов отпадёт.
То есть исследование проводят методом последовательных итераций. Данный метод — это первая итерация, поэтому отсеиваемые виды не ценофобы, а только «кандидаты» в них. Хотя, конечно, это может произвести впечатление подгонки результатов путём отсеивания ненравящихся данных. В какой-то мере от субъективности предохраняет использование двух независимых критериев: (1) распространение отлично от других и (2) нет видов с комплементарным ареалом. Из теории структурализма следует, что ценофобы обычно удовлетворяют обоим критериям, а ценофилы — нет.
Как дополнительный критерий ценофобности вида можно использовать данные о его распределении. Мозаичность распределения свойственна и ценофильным, и ценофобным видам. Но ценофильные виды обильны в определённых ассоциациях = имеют высокую верность (встречаются обычно совместно в определённых ассоциациях, входящих в ту же ассоциацию), а сама ассоциация приурочена к определённым абиотическим условиям. Ценофобные виды имеют малую верность и массовы в местах недавних нарушений. Поскольку нет оснований полагать, что различные ассоциации и сочетания абиотических факторов отличаются частотой нарушений, связь ценофобных видов с абиотическими факторами и другими видами должна быть существенно ниже.
Смотрите также
Понятие исследования
В современном менеджменте исследование является главным фактором успеха, а если выражаться по научному, — главным фактором повышения эффективности управления. Исследование — это не только ...
К популяционной организации политипического вида (на примере рыжей полевки -
clethrionomys glareolus shreb.)
Анализ популяционной организации и динамики численности европейской рыжей полевки
- типичного представителя мегаареальных политипических видов млекопитающих Палеарктики
- подтвердил высказанное на ...
Экономические методы охраны окружающей среды и особенности их использования в России
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...