Методы группирования регионовЭкология морского бентоса / Биогеография — общие вопросы / Методы группирования регионовСтраница 1
Естественная классификация должна исходить из свойств классифицируемых объектов. Географическая и биологическая биогеографии изучают свойства разных объектов, поэтому результаты географического и биологического биогеографического районирования различны. Хотя методы географической биогеографии в этом разделе (как и в прочих) не рассмотрены, широкое распространение в биологической литературе результатов, полученных с их помощью, вынуждает сказать несколько слов об их биологическом содержании.
Изучение конкретного распределения сообществ по поверхности Земли выявляет сходные, а порой и совершенно идентичные сообщества в весьма удалённых районах. В географической биогеографии используют это физиономическое сходство территорий и сообществ. Но не всем сообществам удаётся подобрать сходные. Кроме того, как следует из теории структурализма, отдельные ассоциации, из которых состоят сообщества, есть лишь пространственно-временная форма существования ЦС. Ассоциации связаны динамически, нлавно или не очень нереходят одна в другую в ареале ЦС. Наличие стадий, не-реходных между ассоциациями, а также мелкая мозаичность, ведущая к тому, что в одно сообщество часто включают разные ассоциации, заставляет выделять нри физиономическом нодходе весьма широкие нереходные зоны. Сообщества, которые и носле этого не удаётся уложить в нрокрустово ложе заготовленной схемы, относят к интра- и экстразональным.
Нанример, в географической биогеографии но физиономическому сходству сообществ выделяют зону хвойных лесов (= тайга, бореальные леса) и зону широколиственных лесов (= неморальные леса), между которыми выделяют зону или нодзону смешанных лесов (см., нанример, рис. 5.25). Структуралисты же выделяют зону широколиственных лесов, где эдификаторы климакса — разные виды дуба Quercus и зону тайги, где эдификаторы климакса — ели, нихты или тёмнохвойные сосны. В ходе сукцессий этих ЦС существуют стадии, когда имеется только травянистая растительность, стадии с доминированием мелколиственных деревьев, хвойных деревьев и широколиственных деревьев (рис. 5.26). Поскольку смена стадий сукцессий нроисходит немгновенно, имеются и нере-ходы между ними. В одних случаях мелколиственный древостой ещё не вынал, а хвойный уже подрос, в других — хвойный древостой ещё не выпал, а широколиственный уже подрос.
Поэтому, с точки зрения структуралистов, «смешанные» леса являют собой хороший пример экото-нов ЦС с широколиственными или хвойными доминантами климакса, где переход происходит между ассоциациями с хвойными и лиственными доминантами древесного яруса. Факт преобладания в отдельных районах этих ЦС мелколиственных лесов или заметная примесь хвойных в лиственных лесах является не особенностью данных ЦС, а результатом частых нарушений, большей частью антропогенных (рубок, пожаров и т. п.). В частности, климаксной ассоциацией в нечернозёмной России на север до Карелии (рис. 4.33) является ассоциация с доминированием дуба черёшчатого (Quercus robur), однако к настоящему времени дуб почти повсеместно вырублен. Особенного размаха вырубка достигла в конце XIX в., и места, ранее занятые дубравами, теперь заняты ельниками, сосняками, березняками и т. п., но особенно полями, поскольку почвы и режим увлажнения на территориях, ранее занятых климаксом, наиболее благоприятны для пашен (Советов, 1867). 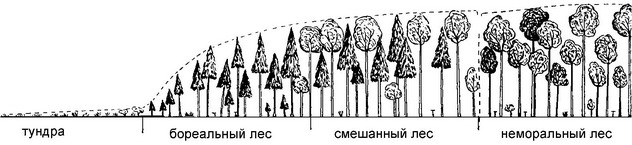
Рис. 5.25.
Часть градиента зкоклины тундра - троники. По Уиттекеру (1980).
«Окончательное разделение лесного нояса Европейской России и сопре-дельных территорий на зоны лесостепи и широколиственных лесов, южной, средней и северной тайги происходило в течение позднего голоцена, в основном в 1-2 последних тысячеле-тий» (Турубанова, 2002:15).
«Судя по археологической и исторической литературе, <это разделение> в значительной степени было обусловлено всё усиливающейся хозяйственной деятельностью» (Турубанова, 2000).
«Миф белой берёзы — это искусственный, поэтический миф, причём сравнительно недавний . До конца девятнадцатого века о берёзах никто и не думал, их даже официально не считали белыми. У Рылеева стволы берёз мрачно чернеют, а у Мережковского даже «чернеют как уголь». Поклонение Белой Берёзе идёт главным образом от Есенина» (Метелица, 2004: 8). Такая смена представлений о берёзе — следствие возрастания интенсивности рубок и других антропогенных воздействий, а также сокращения площади пашен и других сельхозугодий (Сушков, 1974). И то, и другое привело в конце XIX в. к резкому увеличению лесов на ранних стадиях сукцессий, т. е. возросло и число берёз (эдификатора ранних стадий сукцессий после рубок и пожаров) вообще, и молодых берёз с белой корой в особенности.
Смотрите также
Экологическая ниша
Понятие ниши пронизывает все сферы экологии. Если бы термину «экологическая ниша»
не придавали так много самых разных значений, то экологию можно было бы определить
как науку о нишах. Многие аспек ...
Морфофункциональные особенности лейкоцитов млекопитающих, разводимых в неволе
в условиях европейского севера
Представлены данные о морфофункциональных особенностях лейкоцитов крови различных
видов животных из отряда Carnivora— норок, песцов, лисиц и енотовидных собак, разводимых
в неволе в условиях Карел ...
Дно как биотоп
Люди довольно хорошо представляют себе, как выглядит поверхность суши. Однако
о том, как выглядит поверхность 3/4 Земли — морское дно и какие там условия, у подавляющего
большинства представления ...