Островная биогеография и экологияЭкология морского бентоса / Биогеография — общие вопросы / Островная биогеография и экологияСтраница 4
Если же на острове (островах) формируется собственная ЦС (т. е. когда он не может быть заселён существующими ЦС и его возраст достаточно велик) предсказания островной теории становятся особенно неудовлетворительными. Это связано с тем, что островная теория в классическом виде не рассматривает видообразование вовсе. Между тем видообразование, обычно сопровождающее формирование ЦС, может принципиально изменить наблюдаемую картину. Возьмём, например, два крупнейших пресноводных озера мира: Виктория и Верхнее. Оз. Виктория имеет максимальную глубину 80 ми площадь в 68 тыс. км2; оз. Верхнее — 393 м и 82,4 тыс. км2. Оба они существуют примерно одно время — около 10 тыс. лет. Однако, оз. Виктория имеет богатую эндемичную биоту (число эндемичных видов составляет многие сотни, велико и число эндемичных родов), а биота оз. Верхнее ничем не отличается от биот соседних озёр. 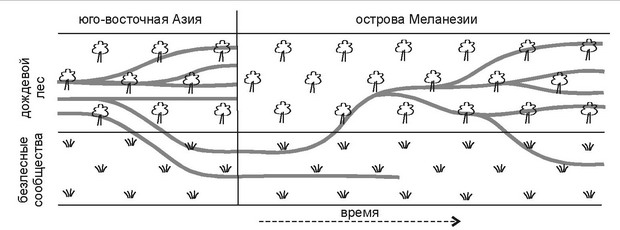
Рис. 5. 34.
Филогения муравьев Меланезии.
Виды безлесных сообществ при заселении островов сначала входят в состав безлесных сообществ, затем лесных, где образуют несколько видов в результате радиации, позднее от них образуются виды, вновь входящие в состав безлесных сообществ. По Wilson (1959).
Островная биота — неслучайная выборка из исходной биоты, не группировка. На островах также имеются ЦС. Отличия в их составе от исходных обусловлены различной способностью видов к расселению. Сукцессии могут останавливаться из-за отсутствия видов, необходимых для прохождения данной стадии (диаспорический субклимакс). Одновременно на островах начинается изменение ЦС так, чтобы наиболее эффективно осуществить использование ресурсов при наличном наборе видов. В первую очередь это приводит к расширению экологических ниш видов, часто в сторону ранних сукцессионных стадий (в случае наземных ЦС — это безлесные сообщества).
Изменение ниши видов отражается в изменении их морфологии. В случае наличия большого количества неиспользуемых ресурсов в результате адаптивной радиации могут возникать потребляющие эти ресурсы виды. В результате таксон проходит как бы цикл: виды поздних сукцессионных стадий при формировании новой ЦС или её изменении сначала входят в состав пионерных сообществ, и лишь затем в состав более зрелых, поздних сукцессионных стадий.
Изучение реальной ситуации в Вест-Индии (Ricklefs, Cox, 1972) показало, что большая часть недавно вселившихся видов действительно несколько расширяют свои ниши, но в целом они приурочены к открытым безлесным сообществам (ранним сукцессионным стадиям), тогда как сравнительно давно вселившиеся виды обитают преимущественно в лесных сообществах (поздних стадиях сукцессии), ниши их сужаются, плотность популяций некоторых из них снижается, других — расширяется (в зависимости от их роли в сообществах). Некоторые виды после вселения сразу занимают узкую нишу и преобразуются в эндемов.
Подобное явление было названо «цикл таксонам». Классическим примером цикла таксона являются результаты исследований R.Wilson (1959), приведённые на рис. 5.34. В действительности, «цикл таксона» является не какой-либо специфической закономерностью эволюции отдельных таксонов, а отражением становления ЦС — т. е. свойством системы более высокого уровня организации, нежели таксон. Поэтому термин «цикл таксона» не просто не информативен, а не корректен. Очевидно, что формирование ЦС может начаться только с ранних стадий, а не, например, с климакса.
Для островных биот по сравнению с материковыми характерны обеднённый видовой состав и упрощённые ЦС, поэтому многие свойства материковых ЦС могут отсутствовать в островных. Степень отличия монотонно возрастает с уменьшением площади, т. е. имеется обратная зависимость между лёгкостью изучения островной ЦС и степенью её отличия от материковых ЦС, изучение которых и является конечной целью большинства работ по исследованию островов. В любом случае, механический перенос выявленных при изучении островных биот закономерностей на материковые невозможен. Более корректным было бы сопоставлять не остров с материком, а ЦС острова (группы островов) с аналогичной ЦС материка. Соответствие теории островной биогеографии фактическому положению вещей тем лучше, чем в большей степени изучаемые острова являются частью крупных ЦС, и тем меньше, чем мельче масштаб, т. е. при переходе с экологического масштаба к биогеографическому точность предсказаний островной теории стремительно снижается. Как вынужден был признать R.J. Whittaker в упомянутой выше «Биогеографии островов», за более, чем 30 летний период интенсивных исследований выяснилось: «a grand unifying theory remains elusive, and perhaps illusory» (Whittaker, 1998: 4). Поэтому, несколько эпатируя, можно сказать, что биогеография островов — удобная модель для изучения островов.
Смотрите также
Основные функции и принципы экологической политики.
Комплексный характер экологических проблем
требует комплексного государственного управления в области охраны окружающей
среды. Ниже перечислим функции такого управления.
* Экологическое
прогно ...
Биогеография
Основная цель экологии состоит в том, чтобы понять те разнообразные факторы,
которые влияют на распределение и обилие животных и растений (Andrewartha, Birch,
1954; Krebs, 1972; Mac-Arthur, 1972). ...
Влияние окружающей среды на живые организмы
Все процессы в биосфере взаимосвязаны. Человечество - лишь незначительная часть биосферы, а человек является л ...