Абиссаль и ультраабиссальЭкология морского бентоса / Биогеография морского бентоса / Абиссаль и ультраабиссаль
Заселение абиссальных глубин океана многоклеточными происходило как минимум трижды: в докембрии, карбоне-перми и кайнозое. Предшествующая фауна гибла (или, по крайней мере, в значительной степени вымирала) в периоды тёплого климата (разбор причин см. Главу 1). Биогеографический ранг ультраабиссали недостаточно установлен, возможно, ультраабиссальные регионы следует рассматривать как отдельные регионы соответствующих абиссальных.
Наиболее детальной является схема районирования абиссали Н.Г. Виноградовой (1956), а ультраабиссали — Г.М. Беляева (1974). Эти схемы были объединены Н.Г. Виноградовой (1977) (рис. 6.14). Она выделила в абиссали три области, каждую из которых подразделила на ряд подобластей и провинций. Ультраабиссальные регионы выделены в ранге провинций в пределах абиссальных регионов. К сожалению, при установлении ранга регионов был использован всё тот же индекс Престона, непригодный для анализа подобных данных, поэтому фактически ранг выделенных регионов не обоснован. Совершенно неясно и соответствие ранга абиссальных и щельфовых регионов: остаётся непонятным, например, чему соответствует область абиссали на щельфе: области или провинции? Нектон в абиссали полностью отсутствует, что, по-видимому, связано с малым количеством ресурсов, делающим активное хищничество энергетически невыгодным. Глубоководные хищники (удильщики Ceratioidei, головоногие моллюски с длинными щупальцами (Mastigoteuthis и др.) используют исключительно стратегию поджидания добычи и не совершают миграций. В ультраабиссали нет даже быстро движущихся организмов (рыб, головоногих моллюсков, десятиногих ракообразных). В абиссали, но не в ультраабиссали широко распространена биолюминесценция. 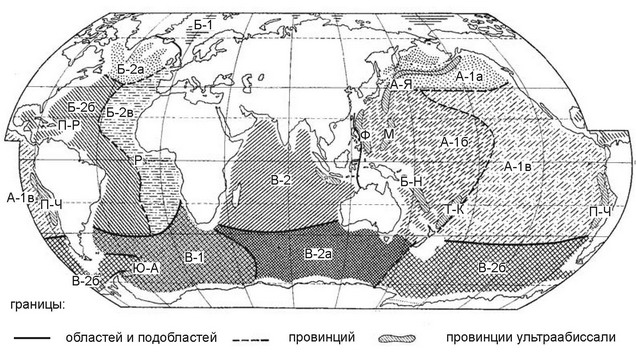
Рис. 6.14.
Районирование абиссали и ультраабиссали Мирового окена Н.Г. Виноградовой (1977).
А — тихоокеанско-североиндийская глубоководная область: А-1 — тихоокеанская подобласть (А-1а — северотихоокеанская, А-1б — западнотихоокеанская, А-1в — восточнотихоокеанская провинции); А-2 — североиндийская подобласть; Б — атлантическая глубоководная область: Б-1 — арктическая подобласть, Б-2 — атлантическая подобласть (провинции: Б-2а — североатлантическая, Б-2б — западпоатлаптическая, Б-2в — восточпоатлаптическая); В — антарктическая глубоководная область: В-1 — антарктическо-атлантическая подобласть, В-2 — антарктическо-индийско-тихоокеанская подобласть (провиднции: В-2а — индийская, В-2б — тихоокеанская). Ультраабиссальные провинции: А-Я — алеутско-японская, Ф — филиппинская, М — марианская, Б-Н — бугенвильско-новогибридская, Т-К — тонго-кермадекская, П-Ч — перуано-чилийская, Я — яванская, П-Р — пуэрториканская, Р — желоба Романш, Ю-А — южноатлантическая.
Рис. 6.15.
Упрощение широтной биогеографической структуры с глубиной на примере брахиопод западной части Тихого океана. Географические элементы фауны: СТ — северо-тихоокеанский, Ян — японский, ЗТ — западнотихооке-анский, ЮА — южноавстралийский, НЗ — новозеландский, Ан — антарктический, точками обозначен элемент фауны, состоящий из видов, распространён-пых по всему Мировому океану. Пределы колебания положения верхней (ВГ) и нижней (ПГ) границ промежуточных вод. По О.П. Зезиной (1973). 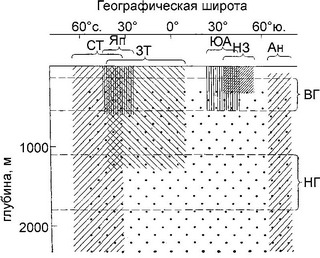
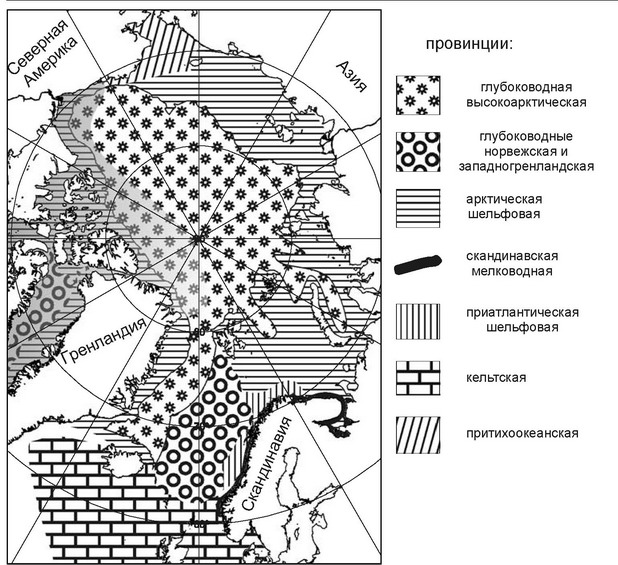
Рис. 6.16.
Схема биогеографического районирования Северного Ледовитого океана но нолихетам. Серым залитої районы, зоогеографическая принадлежность которых установлена исключительно по литературным данным.
Сопоставление биогеографических схем разных вертикальных зон было сделано О.Н. Зезиной (1973), она показала, что с увеличением глубины от шельфа до абиссали происходит уменьшение числа выделяемых регионов (рис. 6.15). Поскольку ультраабиссаль сильно фрагментирована, то здесь вновь происходит увеличения числа регионов.
Смотрите также
Биогеография
Основная цель экологии состоит в том, чтобы понять те разнообразные факторы,
которые влияют на распределение и обилие животных и растений (Andrewartha, Birch,
1954; Krebs, 1972; Mac-Arthur, 1972). ...
Структура сообщества
За исключением краткого рассмотрения биомов в гл. 3, до сих пор мы касались только
экологии особей и популяций. В этой главе речь пойдет об экологии сообществ. Подобно
тому как популяции обладают ...
Экономические механизмы охраны природы
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...