ПопуляцииКниги по екологии / Общая экология / ПопуляцииСтраница 23
В другом случае изменения темпов роста популяций находятся в обратной зависимости от плотности. Разреженная популяция в благоприятных условиях быстро наращивает свою численность, но чем больше становится в ней особей, тем меньше доля очередного приплода, пока скорость воспроизводства не сравняется со смертностью (рис. 126). Тогда темп роста популяции снижается до нуля, а общая численность стабилизируется в соответствии с доступными для популяции ресурсами. Такой характер становления новых популяций свойствен в основном видам, у которых успех в размножении при низком уровне численности не лимитируется обязательностью группового образа жизни, необходимостью встречи полов и другими причинами (например, у мелких партеногенетических ракообразных, растений с апомиксисом и т. д.). Однако у большинства видов наиболее высокий темп роста популяций отмечается лишь при определенной оптимальной плотности. Если популяция сильно разрежена, это затрудняет встречу полов, защиту молодняка, проявление эффекта группы у животных, опыление у растений, поэтому население растет вначале очень медленно. 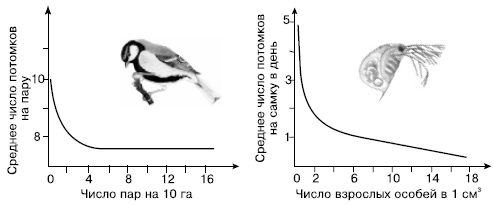
Рис. 126.
Зависимость рождаемости от плотности в лабораторной популяции дафний и в дикой популяции большой синицы (по Ю. Одуму, 1975)
Как известно, появление потомства прежде всего зависит от числа производителей – особей, находящихся в генеративном возрастном состоянии. Вначале рост числа производителей сопровождается медленным ростом численности популяции, затем, в некотором интервале плотностей, зависимость выражена чрезвычайно резко, и даже небольшое увеличение доли производителей вызывает стремительный рост популяции, пока она не достигнет определенного уровня, который в последующем уже не меняется, сколько бы ни увеличивалась репродуктивная часть населения.
Такая зависимость была впервые предсказана французским математиком Верхюльстом в середине XIX века для населения людей, а позднее доказана англичанином Перлом (1925) для популяций животных в среде, где пищевые ресурсы имеют определенный предел восполнимости.
Установление определенного уровня плотности популяций после некоторого периода роста вовсе не означает, что в популяциях больше не происходит количественных изменений. Напротив, любое население всегда динамично и постоянно подвержено колебаниям численности, однако размах всех суточных, сезонных и годовых изменений популяций все же намного меньше теоретически возможного, соответствующего реализации всего биотического потенциала. Колебания численности совершаются с разным размахом вокруг некоторой средней величины, которая и соответствует горизонтальной части кривой на графике роста и стабилизации численности популяции.
Высокий потенциал размножения играет большую роль в выживании видов. Популяции, сведенные к низкому уровню численности, могут быстро восстановиться при благоприятной перемене условий. Некоторые виды только массовым размножением могут противостоять выеданию их различными потребителями или угрозе вытеснения конкурентами. Высокая размножаемость способствует быстрому освоению видом новых пространств.
Однако безграничное размножение таит в себе и большую опасность для любой популяции, так как может привести к быстрому подрыву ресурсов среды, нехватке пищи, убежищ, пространства и т. п., что неминуемо повлечет за собой общее ослабление популяции. Перенаселенность настолько неблагоприятна для любого вида, что в ходе эволюции у разных форм выработались в результате естественного отбора самые разнообразные механизмы, способствующие предотвращению избытка особей и поддержанию определенного уровня плотности популяций.
8.5.7. Динамика ценопопуляций растений
Все размерные и количественные характеристики растений в ценопопуляциях меняются в широком диапазоне.
Меняются такие показатели, как общая численность и плотность, фитомасса, площадь, занятая популяцией, проективное покрытие, возрастной спектр.
У большинства луговых растений колебания численности всходов как в течение сезона, так и по годам составляют от 1–2 до 100-1000 на м2. В степных ценозах в отдельные годы всходы могут не появиться совсем. Массовое их отмирание происходит, например, при наступлении засухи, поедании фитофагами, угнетении взрослыми растениями. Возрастные спектры ценопопуляций разных видов обладают разной степенью динамичности: 1) популяционные волны, (т. е. волны численности) перемещаются постепенно, при этом тип возрастного спектра не меняется и он остается полночленным. Это происходит при регулярном, но относительно небольшом пополнении молодыми особями; 2) популяционные волны перемещаются быстро, возрастной спектр может быть разорванным и неполночленным (рис. 127). Между этими двумя типами динамики различаются всевозможные переходы.
Большая лабильность всех популяционных показателей свойственна видам реактивным, эксплерентам способным очень быстро захватывать освободившиеся площади, но и освобождать их при вытеснении конкурентными видами.
Смотрите также
Основные функции и принципы экологической политики.
Комплексный характер экологических проблем
требует комплексного государственного управления в области охраны окружающей
среды. Ниже перечислим функции такого управления.
* Экологическое
прогно ...
Экономические механизмы охраны природы
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...
Биогеография морского бентоса
в биосфере Земли можно выделить четыре основных типа сравнительно независимых
друг от друга ЦС: морские, пресноводные и наземные. В свою очередь морские ЦС можно
разделить на бентосные, биотоп кот ...