ПопуляцииКниги по екологии / Общая экология / ПопуляцииСтраница 24
Нередко наблюдается однонаправленное необратимое изменение ценопопуляции или ее отдельных локусов от зарождения до зрелости и старения. В конечном счете ценопопуляция или локус на данной площади исчезает. Такой тип динамики называется сукцессивным.
Например, при интенсивной пастбищной нагрузке в ценопопуляциях мятлика однолетнего на залежах уменьшается плотность ценопопуляции, из нее выпадают молодые группы, происходит быстрое общее старение и ценопо-пуляция исчезает. В некоторых случаях сукцессивным типом динамики характеризуются отдельные локусы в пределах ценопопуляции, а сама она в целом сохраняет стабильность. 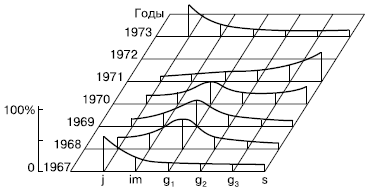
Рис. 127.
Погодичные изменения популяционного потока у полевицы тонкой (по Е. И. Курченко, 1975)
В широколиственных лесах ценопопуляции луковичного растения гусиного лука желтого существуют в виде разобщенных локусов. Этот вид относится к группе эксплерентов, т. е. способен очень быстро захватывать освободившуюся территорию вследствие высокой скорости роста и большой энергии вегетативного размножения. Часто локус занят одним клоном, начало которому дает одна ювенильная особь. Перейдя в имматурное состояние, она начинает размножаться, образуя новые ювенильные особи. Значительная часть ювенильных растений переходит затем в покоящееся состояние, а нормально развивающиеся последовательно проходят все возрастные состояния вплоть до генеративного. В итоге все или почти все особи клона могут перейти в покоящееся состояние. Так завершается развитие локуса. Этот процесс длится 10–25 лет. Но в природе подобное происходит редко, так как даже незначительные нарушения почвы и лесной подстилки роющими животными приводят к пробуждению покоящихся луковичек. Развитие локуса опять начинается с ювенильного возрастного состояния растений, и процесс развития клонов приобретает циклический характер. Так как разные локусы развиваются асинхронно во времени и пространстве, вся ценопопуляция претерпевает флюктуационные изменения. Флюктуации
– это обратимые, разнонаправленные изменения, когда чередуются периоды старения и омоложения ценопопуляции и поколения непрерывно сменяют друг друга. Таким образом популяция сохраняет за собой занятую площадь.
У некоторых луговых растений временное прекращение инспермации и одновременное взросление и старение особей может привести к тому, что возрастной спектр утратит молодую часть, станет разорванным, неполночленным (рис. 128). Когда возобновление восстановится, молодые особи при дальнейшем развитии популяции образуют новую популяционную волну, которая со временем сменит затухающую волну старой части ценопопуляций. Такие волнообразно-флюктуационные колебания численности и возрастной структуры прослежены, например, у луговика дернистого. Высокий динамизм наблюдается и в посевах луговых трав, особенно при действии различных антропогенных факторов, таких, как высокие дозы удобрений, полив, многократное отчуждение. 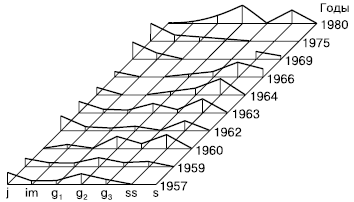
Рис. 128.
Динамика ценопопуляций луговика дернистого на окских лугах (по Л. А. Жуковой, 1986)
8.5.8. Гомеостаз популяций
Поддержание определенной плотности получило название гомеостаза популяций.
В основе способностей популяций к гомеостазу лежат изменения физиологических особенностей, роста, поведения каждой особи в ответ на увеличение или уменьшение числа членов популяции, к которой она принадлежит.
Механизмы популяционного гомеостаза зависят от экологической специфики вида, его подвижности, степени воздействия хищников и паразитов и др. У одних видов они могут проявляться в жесткой форме, приводя к гибели избытка особей, у других – в смягченной, например в понижении плодовитости на основе условных рефлексов. 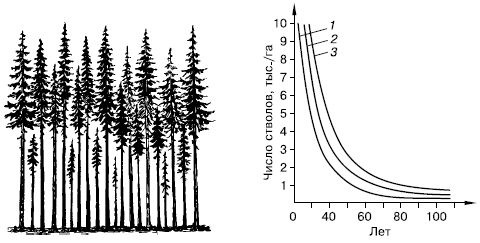
Рис. 129.
Самоизреживание в древесных насаждениях (по Г. Ф. Морозову, 1928):
слева – господствующие и угнетенные деревья в ельнике; справа – ход изреживания стволов с возрастом у сосны (1), березы (2) и ели (3)
К жестким формам внутривидовой конкуренции следует отнести, например, явление самоизреживания
у растений (рис. 129). При большой густоте всходов часть растений неминуемо погибает в результате угнетения физиологически более сильными соседями. Уменьшение числа растений происходит, даже если высеянные семена генетически однородны. В этом случае, по-видимому, имеют значение разница в размерах семян, во времени появления всходов, детали микроокружения. В одном из опытов с клевером Trifolium subterraneum через 84 дня после появления всходов на участке в 1 м2 из 1250 растений осталось 650, причем влияние вредителей было исключено.
У райграса многолетнего основной экологической единицей становится не особь, а побег. Обнаружено, что при разных нормах высева семян, от 6 до 180 кг/га, сначала густота побегов варьирует от 30 до 1070 на 100 см2, но затем во всех случаях становится равной примерно 500, т. е. в более редких посевах появляются новые побеги, а в более густых часть отмирает.
Смотрите также
Особенности природоохранной деятельности в России.
В нашей стране на первом этапе становления хозяйственного
механизма природопользования недостатки административной системы руководства
проявлялись более явно и отчетливо, чем в других странах. ...
Взаимодействие климата и растительности
Климат — главный фактор, определяющий характер растительности. Растения в свою
очередь также в некоторой степени воздействуют на климат. Как климат, так и растительность
оказывают решающее влияние ...
Биологические опасности, связанные с пищей
Научно-технический прогресс сильно повлиял на сферу производства продуктов питания. Технологическая обработка продуктов, консервирование, рафинирование, длительное и неправильное хранение резко снизил ...