ПопуляцииКниги по екологии / Общая экология / ПопуляцииСтраница 29
Общее число жертв, которых может уничтожить один хищник, сначала растет почти пропорционально росту численности потребляемого вида. Это так называемая функциональная реакция хищников на жертвы. Однако она имеет предел, обусловленный физическими возможностями потребителя (рис. 135). После полного насыщения хищники обычно слабо реагируют на жертву. Новые возможности влиять на популяции жертв возникают при росте численности хищников за счет успешного размножения на хорошей кормовой базе. Это явление получило название численной или количественной реакции хищников на рост численности жертв. Количественная реакция хищников всегда запаздывает по сравнению с ходом размножения жертвы, так как является ответом на изменение кормовой базы. 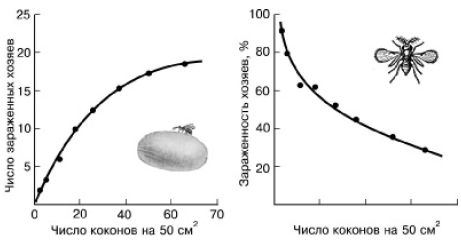
Рис. 135.
Функциональная реакция одиночной самки хальциды Dahlbomi-nus, заражающей коконы пилильщика Neodiprion sertifer (по Дж. Варли и др., 1978): слева – число коконов, пораженных одной самкой паразита, при разной их плотности; справа – зависимость процента пораженных коконов от их плотности при постоянной численности паразита
В лабораторных условиях при совместном содержании хищника и его жертвы отмечаются периодические изменения численности обоих видов, имеющие довольно закономерный характер. В опытах, проведенных нашим отечественным биологом Г. Ф. Гаузе в 30-х годах с пробирочными культурами простейших, было обнаружено следующее. Усиленное размножение хищной инфузории Didinium nasutum закономерно следовало всякий раз за размножением ее жертвы – туфельки Paramecium caudatum. Когда хищник достигал высокой численности, он полностью уничтожал парамеций, после чего наступала гибель самих Didinium, лишенных пищевой базы. Однако, когда в пробирки помещали некоторое количество песчинок, под которыми часть туфелек могла найти недоступные для хищников убежища, после гибели Didinium наступало новое массовое размножение парамеций (рис. 136). В другой серии опытов, когда культивировали разные виды парамеций, а в качестве их пищи – дрожжевые клетки рода Sacharomyces, было получено при дополнительных условиях несколько закономерных циклов колебаний численности дрожжей и туфелек. Механизмы подобных колебаний можно представить следующим образом. 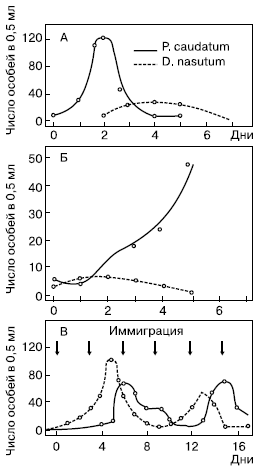
Рис. 136.
Изменение численности инфузорий Paramaecium caudatum (жертва) и Didinium nasutum (хищник) при культивировании в общем сосуде (по Г. Ф. Гаузе, 1934): А – нет убежищ для парамеций; Б – парамеции имеют убежища от хищников; В – с повторным добавлением обоих видов, или иммиграцией
Увеличение численности жертв происходит до тех пор, пока темп выедания их нарастающим количеством хищников превысит скорости размножения. После этого дальнейшая деятельность хищников только подрывает кормовую базу, что ведет к снижению их собственной численности. Это позволяет немногим оставшимся в живых жертвам вновь размножиться и, следовательно, улучшить кормовые условия для хищников. Если последние еще присутствуют в системе, новое увеличение количества жертв способствует новому подъему численности хищников и цикл повторяется снова.
В природных условиях такие закономерные циклические изменения численности двух видов, связанных пищевыми отношениями, выявить трудно, так как на их размножение и гибель влияют взаимодействия со множеством других видов, а также абиотические изменения среды. Однако в ряде случаев и в природной обстановке удается заметить регулярные периодические изменения обилия хищников и их жертв.
Особенно наглядно такие колебания обнаруживаются в обедненных сообществах, где межвидовые связи не столь разнообразны: в тундрах и полярных пустынях, лесах с господством одной породы деревьев, культурных садах и т. п. Циклические изменения численности выявлены, например, у леммингов и их основных потребителей – песцов, массово размножающихся хвое– и листогрызущих насекомых и их паразитов, вредителей яблонь – паутинных клещиков и питающихся ими хищных клещей и у многих других сопряженных пищевыми связями видов. Более детальные исследования показывают, что циклические колебания численности жертв чаще связаны с проявлением внутрипопуляционных механизмов гомеостаза, но и взаимоотношения с хищниками вносят свой вклад в размах этих колебаний.
Таким образом, истребление жертв и поражаемость хозяев зависят от численности врагов, но и обилие последних также определяется численностью объектов их питания.
Паразиты – также мощный фактор сдерживания численности хозяев. Они, как правило, редко вызывают массовую гибель пораженных особей. Но поскольку зараженность паразитами ослабляет организм хозяина, она приводит, как правило, к падению плодовитости или даже полной стерилизации, что имеет следствием снижение численности следующих поколений.
Некоторые паразиты, однако, обязательно приводят хозяина к гибели, что сокращает численность заражаемого ими поколения. Эту группу паразитов называют часто паразитоидами. К ним относятся, например, насекомые, откладывающие яйца в яйца и личинок других насекомых.
Смотрите также
Роль изоферментов лактатдегидрогеназы в адаптациях млекопитающих Карелии
В экстрактах тканей сердца, почек, скелетных мышц, печени, легких, селезенки
у американской норки (Mustela vison L.), песца (Aiopex iagopus), лисицы ( Vuipes
vuipes L.), лесной куницы (Martes mart ...
К популяционной организации политипического вида (на примере рыжей полевки -
clethrionomys glareolus shreb.)
Анализ популяционной организации и динамики численности европейской рыжей полевки
- типичного представителя мегаареальных политипических видов млекопитающих Палеарктики
- подтвердил высказанное на ...
Понятие исследования
В современном менеджменте исследование является главным фактором успеха, а если выражаться по научному, — главным фактором повышения эффективности управления. Исследование — это не только ...