Результаты и обсуждениеЭкспериментальная генетика и физиология / Влияние низкотемпературного закаливания на протеолитическую активность и содержание
фотосинтетических пигментов в листьях проростков озимой пшеницы / Результаты и обсуждениеСтраница 1
Исследования показали, что воздействие температуры 4°С на проростки пшеницы уже через 1 час вызывало небольшое, но достоверное увеличение устойчивости клеток листьев, а к концу четвертых суток она достигала своего максимального значения, сохраняясь в дальнейшем на достигнутом уровне (рис. 1).
Наряду с этим в процессе закаливания у растений наблюдали определенные изменения в активности амидаз и цистеиновых протеиназ. Так, часовое воздействие температуры 4°С вызывало небольшое увеличение амидазной активности, которое затем сменялось ее снижением (на 20% через 5 ч закалки и на 40% - через 24 ч) (рис. 2а). При достижении максимального уровня устойчивости (на 4-е сут. закаливания) амидазная активность проростков пшеницы составляла 40% от максимальной и в дальнейшем она понижалась, достигая значений, характерных для контрольных (не подвергавшихся воздействию холода) растений того же возраста.
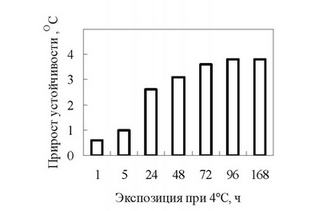
Рис. 1. Влияние низкой закаливающей температуры (4°С) на холодоустойчивость клеток листьев проростков озимой пшеницы с. Московская 39
Активность цистеиновых протеиназ в начальный период охлаждения также увеличивалась, достигая максимума на вторые сутки закаливания (рис. 2б). Дальнейшее воздействие холода вызывало снижение активности данного фермента: на 20% от максимума на 4-е сут. закаливания и на 40% - на 7-е сут. Анализ содержания в листьях фотосинтетических пигментов показал, что холодовое закаливание проростков сопровождается изменением содержания как хлорофиллов, так и каротинои-дов. Причем изменения общего содержания хлорофиллов и количества хлорофиллов в све-тособирающем комплексе (ССК) в целом имели сходную динамику: в течение первых суток закаливания оно снижалось, а затем при достижении максимальной холодоустойчивости постепенно возрастало (рис. 2в, г). Содержание хлорофиллов в фотосистемах (I+II) снижалось через 48 ч от начала закаливания, а затем оно также увеличивалось (рис. 2д). В отличие от этого содержание каротиноидов в течение первых 48 ч действия температуры 4°С сохранялось на одном уровне, после чего несколько возрастало (рис. 2е).
Таким образом, исследования показали, что повышение активности амидаз и цистеиновых протеиназ происходит в начальный период действия на растения пшеницы низкой закаливающей температуры, и оно предшествует росту холодоустойчивости. Это соответствует представлениям о том, что изменение нормальных условий жизнедеятельности сопровождается прежде всего усилением протеолитических процессов (Тарчевский, 2001). Вероятно, протеоли-тические ферменты, контролируя концентрацию белков и пептидов, участвуют в модификации и устранении биополимеров, уже не выполняющих (или выполняющих не в полной мере) в изменившихся температурных условиях присущие им функции, а также обеспечивают клетку (в той или иной степени) мономерными субстратами для синтеза de novo белков, которые участвуют в формировании и поддержании холодоустойчивости клеток.
Помимо этого, в процессе формирования холодоустойчивости по мере увеличения активности амидаз и цистеиновых протеиназ снижается общее содержание хлорофиллов, количество-пигментов ССК, а также хлорофиллов в составе фотосистем (1+11). Из литературы известно, что биосинтез хлорофиллов тесно связан с общим метаболизмом клетки (Литвин, 2000). В частности, метаболиты, образующиеся в ходе обмена белков с участием протеиназ, могут контролировать отдельные звенья синтеза тетрапирро-лов в клетке (а именно этапа фототрансформации протохлорофиллида в хлорофиллид) (Мок-роносов и др., 2006). Поэтому увеличение активности протеиназ в начальный период действия низкой температуры, по-видимому, может способствовать некоторым изменениям (замедлению или полному прекращению) в синтезе пигментов и, как следствие, снижению содержания хлорофиллов. Кроме того, протеиназы хлоропластов, воздействуя на мембранные белки, а также на белки, непосредственно входящие в состав пигмент-белковых комплексов, вызывают целый ряд изменений в составе и конформации белковых молекул, что приводит к нарушению стабильности микроокружения пигментов (Мокроносов и др., 2006).
Смотрите также
Роль изоферментов лактатдегидрогеназы в адаптациях млекопитающих Карелии
В экстрактах тканей сердца, почек, скелетных мышц, печени, легких, селезенки
у американской норки (Mustela vison L.), песца (Aiopex iagopus), лисицы ( Vuipes
vuipes L.), лесной куницы (Martes mart ...
Введение
Исследований, непосредственно посвященных популяционной структуре вида и закономерностям
динамики численности рыжей полевки, немного, к тому же все они носят региональный
характер и основываются н ...
Основные функции и принципы экологической политики.
Комплексный характер экологических проблем
требует комплексного государственного управления в области охраны окружающей
среды. Ниже перечислим функции такого управления.
* Экологическое
прогно ...