Вертикальная зональностьЭкология морского бентоса / Биогеография морского бентоса / Вертикальная зональностьСтраница 4
«Как раньше, так и в настоящее время вертикальное членение донной фауны проводилось по основным структурам дна, . всегда имелись в виду и другие факторы, особенно более резкие градиенты в их распространении» (Шунтов, 2001: 400).
Л.А. Зенкевич (1951: 196) считал 200 м нижней границей распространения донных водорослей, но и это не так, они обнаружены и на вдвое больших глубинах, но, главное, уже на глубине в несколько десятков метров их роль столь незначительна, что выделять на этой основе зону неоправданно.
Другой возможный смысл числа «200 м» — «глубина, глубже которой самые крупные океанские волны уже не могут взмучивать самые мелкие частицы наносов» (Морская геоморфология, 1980). В реальном океане задолго до того, как перестаёт сказываться влияние волн на осадки, перенос осадков волнами становится незаметным на фоне переноса осадков придонными течениями, вызванными иными причинами. Таким образом, и это значение цифры «200 м» незначимо с точки зрения биологии.
Еще одна цитата на тему: «В наши дни глубина 200 м является предельной для широкого экономически выгодного применения существующих технических средств поиска и добычи биологических и минеральных богатств моря» (Баскаков и др., 1987: 17). И это значение не имеет отношения к биологии.
По-видимому, существует только одно сколько-нибудь разумное объяснение широкого использования данного числа: до XIX в., а в некоторых странах и сейчас, расстояния в море (и глубину в том числе) мерили в морских милях и саженях, одна морская сажень (сокращенно fms) = 1/1000 морской мили, 1 морская миля = 1825 м. В схеме Е. Forbes (1856) граница между медиальной и абиссальной зонами проходит по глубине 100 fms. 100 морских саженей — психологически осмысленная величина, тем более, что в то время знания о населении таких глубин были очень незначительны. Эта схема была длительное время широко распространена, и современные схемы вертикальной зональности до некоторой степени являются её развитием. С переходом на метрическую систему мер при-вычные 100 fms было неудобно заменять на 182,5 м, поэтому округлили до 200 м. Ясно, что при таком происхождении величины «200 м», в ней не больше смысла, чем в семи верстах, которые надо пройти, чтоб похлебать киселя. Поэтому использовать 200 м для выде-ления каких-либо биологически осмысленных вертикальных зон более чем странно.
Всякий термин надо определять через свойства объектов, к которым он относится, т. е. биологический термин надо определять через свойства биологических объектов. Поэтому, если уж использовать термины сублитораль, батиаль и абиссаль, то надо вкладывать в них иной, не геоморфологический, а гидробиологический смысл и понимать под ними биогеографические регионы, сменяющие друг друга по глубине. 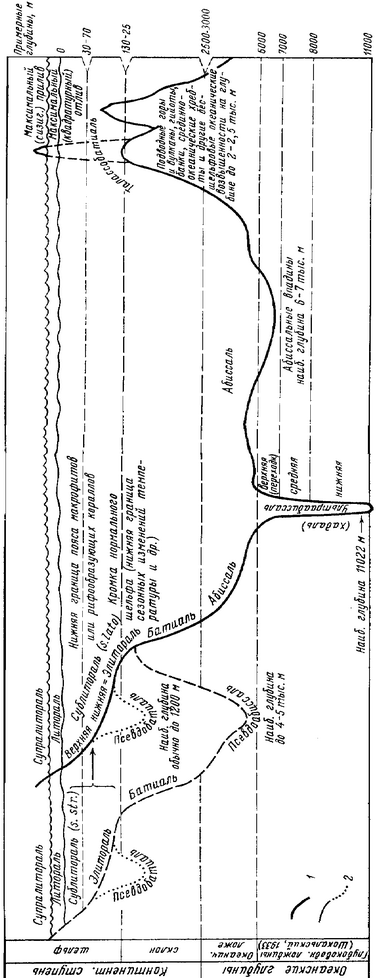
Рис. 6.3.
Схема вертикальной зональности по А.П. Андрияшеву (1979).
Примечание. Нижняя граница распространения рифообразующих кораллов находится на глубине 1-2 км и отнюдь не совпадает с нижней границей пояса макрофитов; по-видимому, имеются в виду герматипные кораллы.
Нельзя не упомянуть схему А.П. Андрияшева (1979) (рис. 6.3). Она является развитием схемы вертикальной зональности 1928 г. (рис. 6.1) Положительными чертами его схемы является разделение сублиторали на верхнюю (= сублитораль 8. 8Іг.) и нижнюю, границу между которыми он проводит по нижней границе пояса макрофитов и рифообразующих кораллов. С этим следует согласиться, уточнив лишь, что не всех рифообразующих кораллов, а герматипных. Дав исторический обзор использования термина «псевдоабиссаль», А.П. Андрияшев указывает на очевидную его непоследовательность* и предлагает полностью изменить его смысл и вводит термин «псевдобатиаль». Он определяет псевдобатиаль как «сравнительно небольшие локальные котловины в пределах шельфа (как, например, в Белом море или на антарктическом шельфе)». Псевдоабиссаль — «большие глубоководные, частично изолированные моря вроде Средиземного или Японского» (Андрияшев, 1979: 123). Судя по приведённым определениям, логическим 
Смотрите также
Платежные инструменты
Платежные инструменты - это правовые средства осуществления переводов денежных средств (наличных и безналичных) с целью совершения платежа.
Порядок выполнения операций с применением платежных инст ...
Морфофункциональные особенности лейкоцитов млекопитающих, разводимых в неволе
в условиях европейского севера
Представлены данные о морфофункциональных особенностях лейкоцитов крови различных
видов животных из отряда Carnivora— норок, песцов, лисиц и енотовидных собак, разводимых
в неволе в условиях Карел ...
Влияние низкотемпературного закаливания на протеолитическую активность и содержание
фотосинтетических пигментов в листьях проростков озимой пшеницы
На проростках озимой пшеницы (Тп11сит aestivum 1.) изучена динамика активности
амидаз, цистеиновых про-теиназ, а также содержания фотосинтетических пигментов при
холодовом (4°С) закаливании. Показ ...